Мозаичность растительности на старых участках поймы малой реки

Особенно актуален такой подход при изучении пойм малых рек.
В этих поймах физиономически и экотопически однородные контуры растительности обычно имеют такие небольшие размеры, что должны рассматриваться не как сообщества, а как фрагменты сообществ -по крайней мере, потому что ни один такой фрагмент не способен полностью вместить элементарную демографическую единицу (ЭДЕ) какого-либо эдификаторного древесного вида.
Площадь, необходимую для размещения ЭДЕ, может иметь только объединение нескольких контуров растительности, входящих в состав пойменного массива. Такое объединение не соответствует фитоценозу в традиционном понимании, и разместившаяся в нем ЭДЕ древесного вида не соответствует классической ценопопуляции. Если это объединение “собрано” из смежных контуров, то оно включает разные элементы или даже формы рельефа, то есть является внутренне резко неоднородным по эдафическим условиям и по видовому составу растительности.
Причем его неоднородность создана не жизнедеятельностью биоты, в отличие от парцеллярной мозаики, но абиотическими факторами - эрозионноаккумулятивной деятельностью русла и поемностью. Если же в такое объединение включены только однотипные по экологическому режиму и составу растительности контуры, то оно не может быть территориально целостным и компактным, так как положительные и отрицательные формы рельефа в пойменном массиве регулярно чередуются.
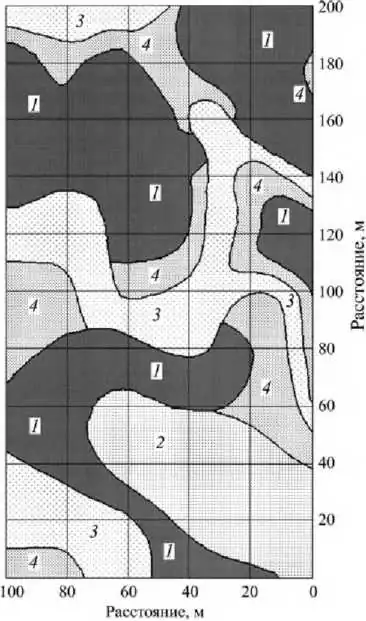
Мозаичность растительности на старых участках поймы малой реки (р. Нерусса) (Евстигнеев, Браславская, 1999).
Условные обозначения: 1 - широколиственный лес, ассоциация Aceri campestris-Quercetum roboris, на краткопоемных верхних участках пойменных массивов, 2 - широколиственный лес, ассоциация Ficario-Ulmetum, на среднепоемных склоновых участках массивов, 3 - широколиственный лес, ассоциация Circaeo-Alnetum, на среднепоемных краевых участках массивов, 4 - травяная растительность, класс Phragmiti-Magnocaricetea, в долгопоемных ложбинах.