Теллермановский массив

Теллермановский массив имеет давнюю историю природопользования, особенность которой - длительное сохранение лесной территории в сочетании с многократными выборочными и сплошными рубками и посадками дуба.
Этот массив был отнесен к корабельным рощам в петровские времена и до 1904 года в нем сохранялись отдельные деревья дуба возрастом более 300 лет с клеймами Петра I (Корнаковский, 1904; Краснопольский, 1928). Изучение лесоустроительных материалов с 1860 года показало, что старые дубовые насаждения, существовавшие до середины (конца) XX века, возникли на месте выборочных рубок дуба на кораблестроение (с начала XVIII века) и вырубок спутников дуба местным населением. При умеренной вырубке дуба в сочетании с его посадкой и постоянным уходом за посадками, дуб сохранял господство в первом ярусе; спутники дуба - липа, клен, ильмовые, - возобновляясь как семенным, так и порослевым путем, постепенно врастали в первый ярус. В результате формировались полидоминантные насаждения. На участках, где проводилась сплошная рубка с нарушением напочвенного покрова, развивались осиновые насаждения. В конце XIX-XX веках выборочные рубки сменились сплошными, целенаправленное выращивание дуба почти прекратилось и в результате образовались широколиственные леса, где дуб встречается единично, а доминируют ясень, липа, реже клен остролистный, ильм.
В пределах экологического профиля выделено 12 экотопов, которые различаются по положению в рельефе, почвам и доминантам в первом подъярусе древесного яруса.
Фитохоры этих экотопов различаются по почвам и доминантам древесного яруса и травяного покрова. Сравнение долей участия видов в разных подъярусах яруса А и яруса В показывает, что по мере развития насаждения из него полностью выпадет дуб и возрастает число видов деревьев в первом подъярусе яруса А. Все насаждения, которые на время их описания (Петров, 1956; Елагин, 1963) различались доминированием видов деревьев, в результате смены поколений окажутся практически одинаковыми.Определение эколого-ценотической структуры фитохор, занимающих разные экотопы, показало, что их можно объединить в три группы.
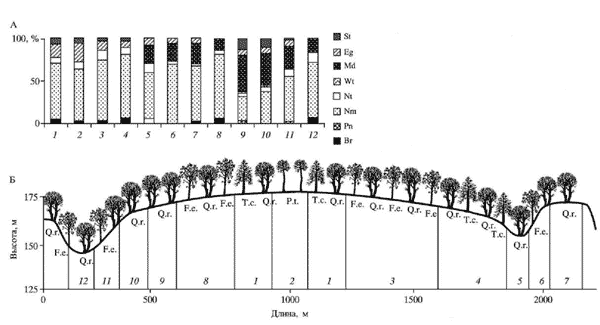
Схема катены и распределение лесного покрова в эрозионном типе ландшафта (Теллермановское лесничество Воронежской области).
А - эколого-ценотические спектры сообществ, Б - размещение экотопов по рельефу.
Обозначение видов деревьев и эколого-ценотических групп.
Экотопы: сведения об экотопах 1-12 приведены в таблице.
Растительность и почвы разных экотопов на экологическом профиле через Теллермановский массив (Воронежская область) широколиственных лесов (Петров, 1956; Елагин, 1963)
|
№ экотопа |
Положение в рельефе |
Ассоциации по доминантной классификации |
Почвы |
|
1 |
Ровные участки широких плато |
Липо-дубняк осоково-снытевый |
Темно-серые лесные хорошо оструктуренные слабооподзоленные тяжелые суглинки |
|
2 |
Небольшие депрессии в пределах широких плато |
Осинник волосисто осоковый |
Серые лесные слабо оструктуренные средне-оподзоленные суглинки |
|
3 |
Верхняя часть пологих склонов (1-3°) северной экспозиции |
Ясене-дубняк снытевый |
Темно-серые лесные хорошо оструктуренные слабооподзоленные тяжелые суглинки |
|
4 |
Нижняя часть пологих склонов (1-3°) северной экспозиции |
Липо-дубняк пролесниковый |
Темно-серые лесные хорошо оструктуренные слабооподзоленные средние суглинки |
|
5 |
Днища плоских балок |
Липо-дубняк крапивный |
Темно-серые лесные слабооструктуренные слабооподзоленные на аллювиально-делювиальных суглинках |
|
6 |
Верхняя часть покатых склонов (5-7°) узких меж-овражных мысов южной экспозиции |
Ясене-дубняк ландышево- осоковый |
Серые лесные слабооструктуренные средне-оподзоленные легкие суглинки |
|
7 |
Плоские вершины узких межовражных мысов |
Ясене-дубняк волосисто осоковый |
Серые лесные слабооструктуренные средне-оподзоленные средние суглинки |
|
8 |
Верхняя часть пологих склонов (1-3°) южной экспозиции |
Ясене-дубняк осоково- снытевый |
Темно-серые лесные хорошо оструктуренные слабооподзоленные средние суглинки |
|
9 |
Средняя часть пологих склонов (1-3°) южной экспозиции |
Дубняк солонцовый |
Оподзоленно-осолоделые солонцы на глинах |
|
10 |
Бровки крутых склонов балок южной экспозиции |
Дубняк кустарниковый |
Оподзоленно-осолоделые солончаково-солонцовые почвы на глинах |
|
11 |
Верхняя часть крутых склонов (10-15°) балок южной экспозиции |
Ясене-дубняк крутосклоновый |
Коричнево-серые на карбонатном суглинке |
|
12 |
Нижняя часть крутых склонов (10-15°) балок южной экспозиции |
Ясене-дубняк овражный |
Серые лесные маломощные на карбонантных суглинках |
1. Фитохоры (дубняк солонцовый и дубняк кустарниковый), занимающие экотопы с солонцовым типом почвообразования и контактирующие с травяными фитохорами на солонцовых полянах. Здесь приблизительно в равных количествах в травяном покрове участвуют неморальные (лесные и опушечные) и луговые совместно со степными видами. Эти фитохоры характеризуются наибольшим видовым богатством в целом и наибольшим, среди всех остальных фитохор, числом видов деревьев, кустарников и, особенно, трав.
Для этой группы фитохор характерен специфический ЭЦК, который не был описан ранее. По преобладанию эколого-ценотических групп он назван неморально-опушечно-лугово-степным (Nm-Eg-Md+St).
2. Фитохоры (ясене-дубняк ландышево-осоковый и ясене-дубняк волосистоосоковый), занимающие экотопы покатых склонов южной экспозиции или узких межовражных мысов со смытыми почвами, верхние горизонты которых сильно пересыхают летом (Петров, 1956). В травяном покрове сообществ этих фитохор в минимуме представлены нитрофильные виды, что связано с большой сухостью их местообитаний.
3. Все остальные фитохоры занимают экотопы плато пологих и крутых склонов и плоских днищ балок с темно-серыми, серыми лесными почвами разной степени оструктуренности. В травяном покрове сообществ этих фитохор 60% и более приходится на неморальные лесные виды, заметное участие принимают неморальные опушечные и нитрофильные виды.

По эколого-ценотической структуре сообщества второй и третьей групп фитохор слабо различаются и относятся к неморальному эколого-ценотическому комплексу, который подробно описан в разделе. Анализ распределения экологических характеристик растительных сообществ экотопов в осях экологических факторов по увлажнению и трофности, а также увлажнению и кислотности почв показал, что все описания растительных сообществ разделились на две части.
Одну часть образовали описания растительных сообществ первой группы фитохор, которые имеют более высокие баллы трофности и кислотности, и более низкие - увлажнения. Такие экологические характеристики имеют остепненные дубравы юга зоны широколиственных лесов на границе леса и степей, и байрачные дубравы степной зоны (Киселева, 1994; Оценка..., 2000).
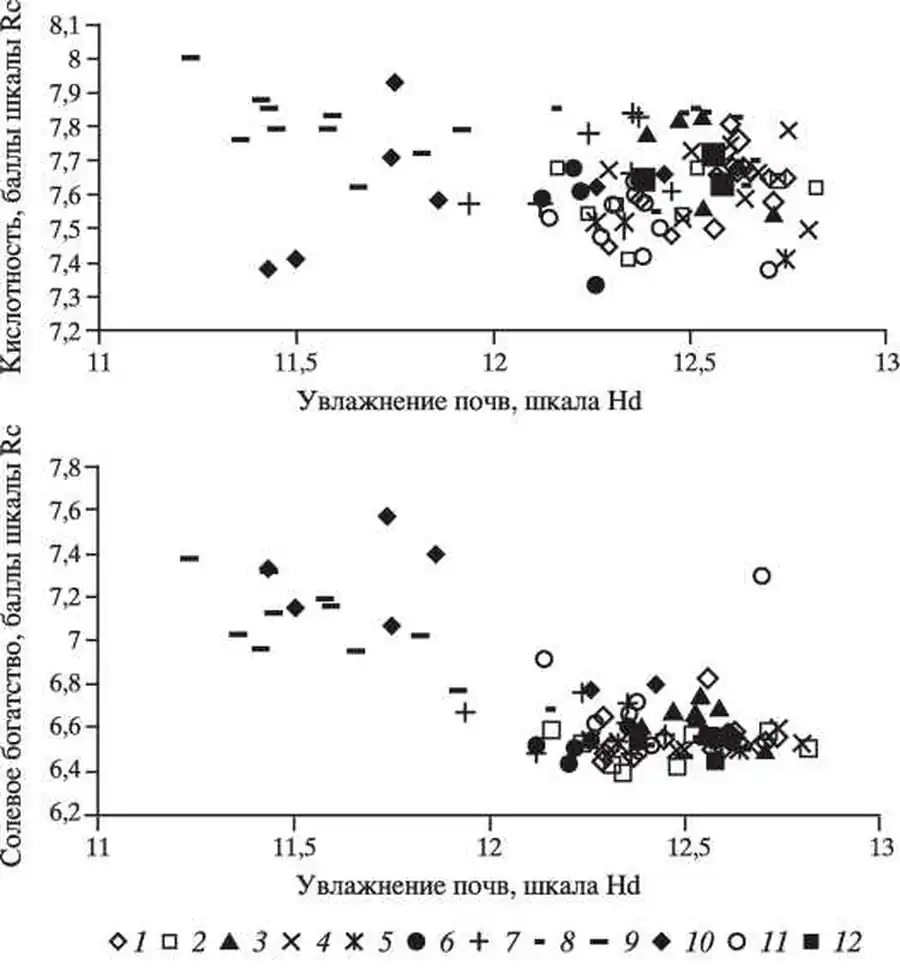
Экологическое пространство катены (Теллермановское лесничество Воронежской области).
Вторая часть описаний относится к растительным сообществам второй и третьей группы фитохор. Эти сообщества имеют более низкие баллы трофности и кислотности и более высокие - увлажнения. Такие экологические характеристики обычны для широколиственных лесов на темно-серых и серых лесных почвах в пределах большей части зоны (Оценка..., 2000).
Таким образом, эколого-ценотический и экологический анализ растительного покрова экологического профиля в Теллермановском массиве широколиственных лесов позволяет предположить, что потенциально растительность катен на юге зоны широколиственных лесов была представлена растительными сообществами двух ассоциаций. Сравнение видового состава подроста и травяного покрова исследованных сообществ позволяет предположить, что потенциальная растительность неполночленной катены в южной части зоны широколиственных лесов представлена сообществами, относящимися к разным порядкам класса Querco-Fagetea: ксерофильных лесов на почвах солонцового типа и мезофильных лесов на серых и на темно-серых лесных почвах ассоциации Aceri campestris-Quercetum. В доагрикультурные времена, в связи с большей оводненностью водотоков, такие катены могли быть полночленными и на днищах глубоких балок растительность была представлена ассоциацией, относящейся, скорее всего, к союзу Alno-Padion (Knapp, 1942) того же класса.