Распространение мамонта в Евразии в позднем плейстоцене

В позднем плейстоцене область распространения мамонта в Евразии сопоставима с Палеарктикой и простиралась на севере от о. Большевик в архипелаге Северная Земля, на юге до левобережья среднего течения Янцзы и Леванта, на западе от Британских островов и Пиренейского полуострова, на востоке до о. Врангеля, Чукотки, Камчатки, Приморья, Монголии, Кореи, о. Сахалин и о. Хоккайдо (Sandergen, 1950; Флеров и др., 1955; Цейнер, 1963; Matthes, 1962; Геологическое..., 1968; Кальке, 1976; Каталог..., 1981; Деревянко, 1983; Аверьянов и др., 1995 и др.).
В Северной Евразии местонахождения остатков мамонта располагаются практически всюду, кроме Средней Азии и части Казахстана.
По радиоуглеродным данным, полученным в последние десятилетия, на пространстве от Днестра и Финляндии до Гыдана, Таймыра и ряда других районов крайнего севера материковой Евразии мамонт дожил до начала голоцена. Наиболее поздние находки его остатков на материковой части Евразии имеют абсолютный возраст около 9 000 л.н., на Ямале - 7200 л.н., на острове Врангеля - около 3900-3700 л.н. (Васильчук и др., 1983; Вартанян и др., 1992; Аверьянов и др., 1995; Сулержицкий, 1995; Kuznetsova et al., 2001).
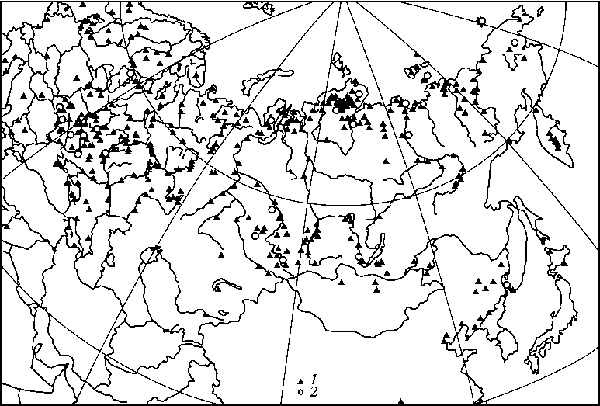
Изменение ареала мамонта в позднем плейстоцене и голоцене:
1 - находки остатков мамонта в позднем плейстоцене; 2 - находки остатков мамонта в голоцене
Наличие остатков мамонтов в наиболее поздние времена в подавляющем большинстве палеореконструкций расценивается как свидетельство крайне суровых (перигляциальных) природных условий (Верещагин, Громов, 1977; Величко и др., 1992). В качестве аргумента приводятся морфологические особенности мамонта: мощный меховой покров, обросший шерстью хобот, уши в 5-6 раз меньшие по площади, чем у азиатского слона, и в 10-12 раз меньшие, чем у африканского слона, короткий волосатый хвост, обильные жировые отложения под кожей (Верещагин, 1971, 1979; Шило, 2001). Однако, представлению о крайне суровых условиях территорий, где обитали мамонты, противоречит ряд данных. Так, в настоящее время в пределах южной границы ареала мамонта, в среднем течение Янцзы, сохраняется теплолюбивый вид - китайский аллигатор (который более нигде не встречается). В Леванте, поблизости от южной границы распространения мамонта, последние нильские крокодилы были уничтожены в 30-е годы XX в., но еще сохраняется реликтовый, изолированный участок ареала африканской трехкоготной черепахи Trionyx triunguis (распространенной на большей части африканского континента). Во Флориде и по нижнему течению Миссисипи по-прежнему обитает американский аллигатор (Иорданский, 1969; Дроздов, 1969). Все эти виды никак не могли бы сосуществовать с мамонтом в условиях холодного и сухого климата, который моделируется в большинстве палеореконструкций для всего ареала мамонта.
У северной границы ареала, на Новосибирских островах, вместе с мамонтом существовала богатая фауна (Черский, 1891), о чем уже сказано выше.
Материалы по питанию мамонта позволяют представить облик растительного покрова. Летом мамонты питались преимущественно травой (луговыми злаками, осоками, пушицей), молодыми побегами и корой деревьев и кустарников (ив, берез, ольхи, чозении, лиственницы); зимой - подсохшей травой, побегами лиственных кустарников и хвойных деревьев (лиственницы, сосны, пихты) (Гетчинсон, 1899; Горлова, 1982; Украинцева, 1996). Суточная норма пищи крупного мамонта могла достигать 3,5-4,0 ц.

Изучение особенностей питания мамонта у северной границы ареала позволило заключить, что в позднем плейстоцене граница распространения древесной растительности в северной Евразии проходила значительно севернее современной. Главной причиной этого было более северное, чем сейчас, простирание евразийской суши, обусловленное наиболее мощной за плейстоцен регрессией Мирового океана. Соответственно и климат на крайнем севере Сибири в то время был более благоприятный для различных крупных травоядных животных, чем в настоящее время, о чем писал еще И.Д. Черский (1891). Значительно севернее была граница криогенных вюрмских степей и лесостепей, тогда как в настоящее время от этой гигантской гиперзоны сохранились на северо-востоке Азии вплоть до Гренландии лишь очень ограниченные по площади реликтовые участки степной растительности (Юрцев, 1976).
Разумеется, по громадной меридиональной протяженности эта гиперзона не была однородной. В силу радиационной зональности и действия региональных факторов, климатические условия на различных ее территориях были различны. Общим же для нее было абсолютное господство открытых и полуоткрытых ландшафтов и наличие лесных сообществ преимущественно в поймах рек (Тугаринов, 1929; Формозов, 1969 и др.).
В течение древнего и раннего голоцена мамонт исчезает почти на всей территории Северной Евразии, о чем можно судить по катастрофическому сокращению числа находок. При этом каких-либо закономерных изменений границ ареала по сравнению с поздним плейстоценом не отмечено. Остатки костей мамонта найдены в тех же районах, где он обитал в конце плейстоцена. В среднем и позднем голоцене остатков мамонта на материковой части Евразии не обнаружено. Наиболее поздние находки - остатки 24 мамонтов с острова Врангеля, датируемые от 7710 до 3730 л.н. (Вартанян и др., 1992; Аверьянов и др., 1995; Сулержицкий, 1995).