Эксперименты с трехвидовыми древостоями

Эксперименты с трехвидовыми древостоями. В настоящее время в литературе активно обсуждается проблема возобновления дуба и формирования естественных насаждений с преобладанием дуба. Была проведена серия модельных экспериментов, в которых меняли параметры экзогенного отчуждения особей клена (как самого теневыносливого вида) в разных возрастных состояниях.
На рисунке представлены результаты одного из экспериментов, в котором были заданы следующие правила периодического (на каждом шаге) отчуждения особей: у березы и дуба удаляли 5% особей всех онтогенетических состояний (имитация случайного отпада деревьев), а у клена -75% имматурных особей (имитация выедания молодых особей клена животными). Кроме графика изменения площади генеративных особей всех моделируемых видов по шагам моделирования представлены аналогичные зависимости для площадей, занимаемых особями разных онтогенетических состояний каждого вида.
Рассмотрим последовательно изменение структуры древесной синузии по шагам моделирования. Вначале территорию очень быстро захватывает береза. Она полностью доминирует в древесной синузии и занимает более 90% территории. Эта ситуация характеризует начальный этап сукцессий в березняках, развивающихся на заброшенных пашнях в условиях близкого нахождения многовидовых широколиственных лесов и возможности постоянного заноса зачатков.
Картина резко меняется после 11-го шага моделирования, когда начинается естественный распад березняка, практически завершающийся к 14-му шагу. К этому времени уже хорошо развит подрост дуба, который начал активно развиваться после 10-го шага с момента начала перехода берез в сенильное состояние. Если бы моделировалась древесная синузия без экзогенных воздействий, то на его месте был бы клен. Он через 2-3 шага занял бы доминирующее положение и не терял его до конца исследуемого периода. Однако периодические отчуждения поддерживают численность клена на допустимом уровне, тем самым искусственно формируя многовидовой древостой. Это позволяет развиваться популяции дуба. В результате уже к 35-му шагу формируется структура, которая практически дальше не меняется. В качестве примера стационарной структуры рассмотрим расположение деревьев березы, дуба и клена 1-, 2- и 3-го ярусов на 75-м шаге моделирования. В полученном древостое дуб занимает ведущее положение и контролирует 35-40% территории, а клен и береза - по 10%. В сумме все виды в генеративном состоянии контролируют в среднем около 60% территории.
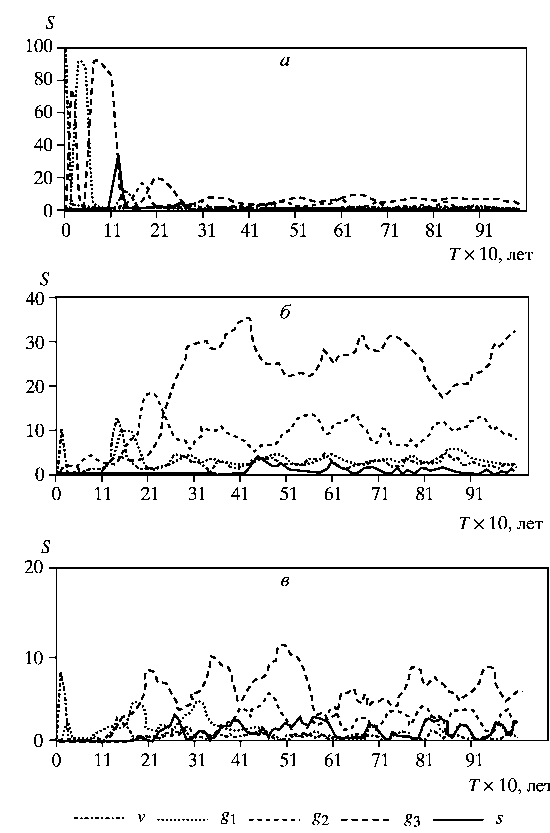
Изменение долей площадей виргинильных, молодых, средневозрастных, старых генеративных и сенильных особей березы (а), дуба (б) и клена (в) в процессе моделирования
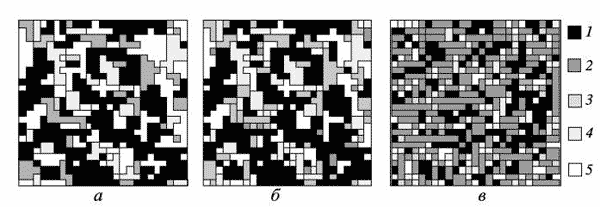
Структура первого (а), второго (б) и третьего (в) ярусов трехвидового лесного сообщества на 75-м шаге моделирования 1 - дуб; 2 - клен; 3 - береза; 4 - смесь видов; 5 - окна

Размещение окон (белые участки) в пологе 2-го яруса на 60-м, 75-м и 90-м шагах моделирования
Случайное сочетание факторов, действующих на древостой, приводит к образованию окон размером от 25 до 450 м2, что позволяет нормально развиваться даже светолюбивой березе. На рисунке представлены структуры древесного яруса на уровне 2-го яруса для шагов моделирования 60, 75 и 90. Конфигурация и размещение окон меняются, но общее их число и размеры остаются практически постоянными. Ниже показано распределение окон разных размеров в перерасчете на 1 га в трехвидовом разновозрастном древостое, достигшем стационарного состояния: среднее число окон площадью около 25 м2 составляет 15,2 шт.; 50-100 м2 - 7,6 шт.; 100-200 м2 - 3,9 шт.; 200-450 м2 - 1,2 шт.
Согласно классификации, данной А.А. Чистяковой (Восточноевропейские..., 1994), окна до 200 м2 отнесены к малым окнам, от 200 до 600 м2 - к средним, более 600 м2 - к большим. При этом подчеркивается, что большие окна образуются обычно в результате экзогенных по отношению к популяционной жизни деревьев воздействий катастрофического характера. В настоящих вычислительных экспериментах не ставилась задача исследования подобных воздействий. Размеры окон, полученные в процессе моделирования, соответствуют реально наблюдаемым в ненарушенных лесах.
Структуры, возникшие на этом этапе вычислительных экспериментов в процессе моделирования разновозрастной многовидовой древесной синузии, в основном правильно воспроизводят особенности мозаично-ярусной организации ненарушенных лесных ценозов. Это становится очевидным при сравнении результатов моделирования с общими чертами структуры таких ценозов, полученными из литературных источников, и с конкретными параметрами одного из ценозов в разновозрастных широколиственных лесах Калужских засек (Смирнова и др., 1991; Восточноевропейские..., 1994).
Проведенные модельные эксперименты показывают возможности модели не только адекватно отразить природные объекты, но и проанализировать влияние эндогенной и экзогенной регуляции на устойчивость организации древесной синузии и лесных сообществ в целом.

Данную имитационную биоэкологическую модель развития многовидового разновозрастного лесного ценоза можно считать базовой. Основная ее особенность - это представление пространства набором ячеек, каждая из которых, с точки зрения расчета доступной ФАР, является полупрозрачным объектом. Изменяя размеры ячейки можно получить целый ряд моделей, имитирующих развитие насаждений на разных пространственных уровнях и зависящих от задач, поставленных перед исследователями (Чумаченко, 1992). Алгоритм расчета доступной ФАР при этом меняется незначительно. Уменьшение размера ячеек позволяет более подробно моделировать крону деревьев и конкуренцию за пространство.
Так, например, уменьшив размеры ячейки до 3-5 м2, можно исследовать динамику древесной синузии с учетом ветровально-почвенного комплекса. В этом варианте алгоритм модели не потребует значительных доработок по сравнению с базовой моделью и позволит изучить влияние условий микроместообитания на возобновление видов в конкретных условиях произрастания.
Дальнейшее уменьшение размеров ячейки до 0,1-0,25 м2 по площади и 0,5-2,0 м по высоте позволяет производить достаточно тонкие расчеты световых характеристик в древесной синузии и внутри кроны и рассчитать пространственное распределение фракций фитомассы древесных видов в лесном ценозе (Носова и др., 1995).
Увеличение размера ячеек до размеров, превышающих площадь кроны взрослого дерева, не позволяет моделировать пространственное размещение отдельных особей - особи не имеют точной пространственной привязки, а считаются случайно расположенными в пределах ячейки. Такой подход позволяет моделировать большие участки, но основные характеристики получаются усредненными по когортам отдельных древесных видов.
Если увеличить размеры ячеек в 3-5 раз (до 100-200 м2), можно моделировать динамику крупных лесных массивов площадью в сотни и тысячи гектаров. При этом можно моделировать неоднородности условий местопроизрастания, зависимость разноса семян как функцию расстояния до генеративных особей.
Представленная модель послужила основой для разработки целой серии моделей, одна из которых представлена в разделе. Учет пространственной, видовой, возрастной структуры конкретных лесных насаждений и возможность моделирования разнообразных антропогенных воздействий позволили разработать прикладную модель динамики лесных насаждений на локальном уровне.