Детерминация нейродермальных мышечных тканей
Факты такого рода и заставляют нас не считать сообщения о возможности превращения глиоцитов в миоциты и миосимпласты заведомо ошибочными.
Но мы допускаем, что часть таких сообщений преждевременна, а более надежные факты не имеют пока теоретического объяснения. Верно, что связь детерминации и дифференцировки не всегда однозначна, но, конечно, это не указывает на то, что дифференцировка может быть любой.
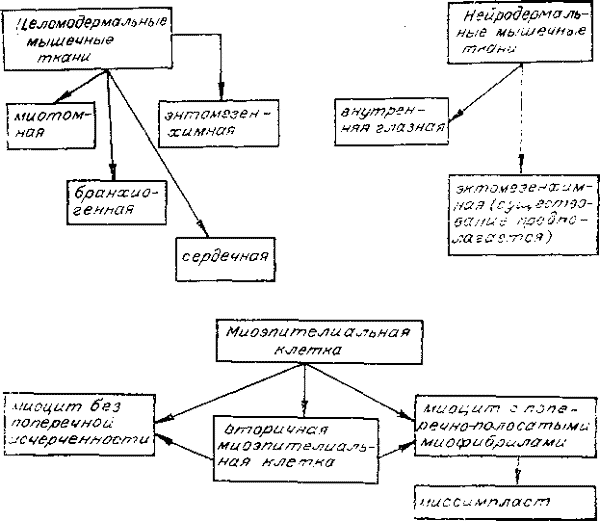
Генетическая классификация мышечных тканей и пути типичной эволюции сократимых элементов.
Что касается детерминации нейродермальных мышечных тканей, то в изученных случаях [Хлопин, 1946] она оказалась специфичной. При утрате мышечного фенотипа клетки в культуре обнаружили рост, сходный с «узорами» роста глиоцитов. Способ взаимодействия определенной клетки с другими задается ее детерминацией, и потому даже при утрате дифференцировки клетки в культуре взаимодействуют определенным образом. Качество отдельной клетки часто нельзя определить, но их «коллектив» образует стандартную для какой-либо ткани картину (что мы и обозначили как узор, чтобы не употреблять еще «не обрусевшего» слова «паттерн»).
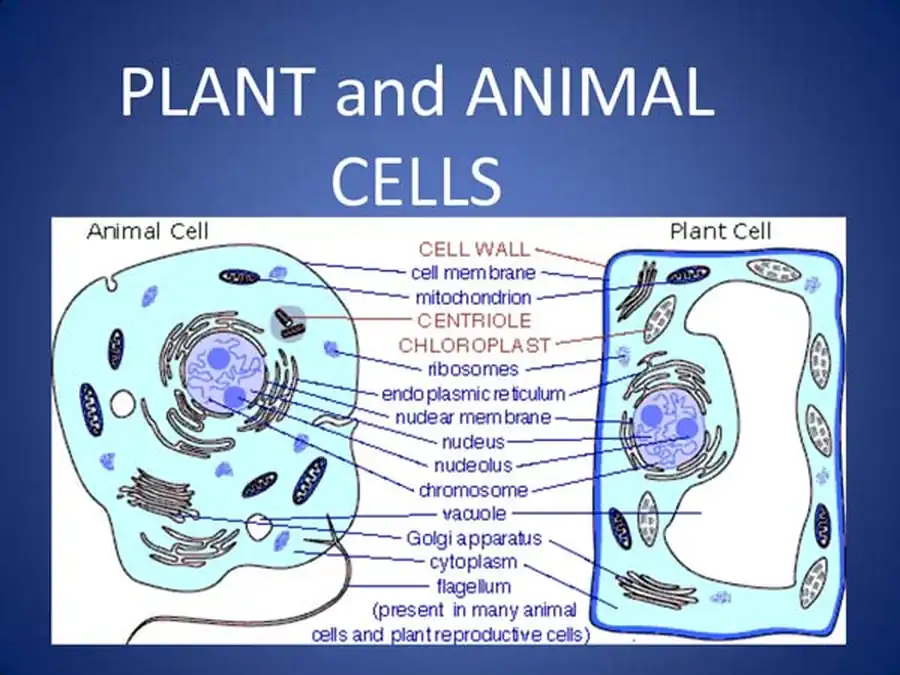
В заключение отметим, что по морфофункциональным критериям у высших позвоночных мы можем выделить четыре вида мышечных тканей: поперечно-полосатую миосимпластическую, сердечную, гладкую и пигментную. Первые три ткани общепризнанны. Пигментной мышечной тканью мы называем радужинную часть сетчатки в той ее части, где миоэпителиальные клетки, содержащие пигмент, преобладают.
По генетическим признакам мышечные ткани делятся на целомические (мезодермальные) и нейродермальные. Устойчивых корреляций между двумя этими классификациями нет. Мы склонны объяснять это тем, что миоэпителиальная клетка является исходной формой для построения любых мышечных тканей. И независимо от ее принадлежности неспециализированной ткани покрова или целомодермы основные пути ее эволюции заданы в первичной дифференцировке и оказываются близкими для разных мышечных тканей.