Индукция нервной пластинки дорсальной губой бластопора
Следовательно, в онтогенезе позвоночных нервная пластинка образуется там, где в филогенезе замкнулась середина первичного рта, и ее образование рекапитулирует перемещение археостома с заднего конца тела на середину.
Индукция нервной пластинки дорсальной губой бластопора (эквивалент смещения бластопора) интенсивно изучалась экспериментальными методами. Но попытки понять процесс как таковой, без привлечения филогенетических соображений, не привели к полному успеху. Из полученных данных для нас наиболее интересно то, что в индукции важное, если не решающее значение имеет процесс дезинтеграции эктодермы [Токин, 1977; Saxen, Toivonen, 1962]. Эффективны как механическое повреждение, так и гибель части клеток и даже просто ослабление связей между клетками.
Можно предполагать, что временное ослабление связей клеток как раз и моделирует образование бластопора на новом месте. А эквивалент бластопора индуцирует, в свою очередь, циркумбластопоральное нервное сплетение, превращающееся в нервную пластинку. Разделение бластопора на рот и анус не воспроизводится не только потому, что бластопор представлен лишь в виде эквивалента, а скорее потому, что это у позвоночных уже невозможно: их рот возник из пары жаберных щелей [Шмальгаузен, 1937] и с бластопором никак не связан.
Нервно-кишечный канал позвоночных отражает действительную филогенетическую связь ЦНС предков позвоночных сзади с анусом. На переднем конце эта связь не сохранилась (у асцидий она есть), так как вторичный рот у позвоночных заместился «третичным». Древние соотношения отчасти отражают отношения переднего края нервной пластинки и обонятельных плакод, которые первично располагались у входа в пищеварительный тракт.
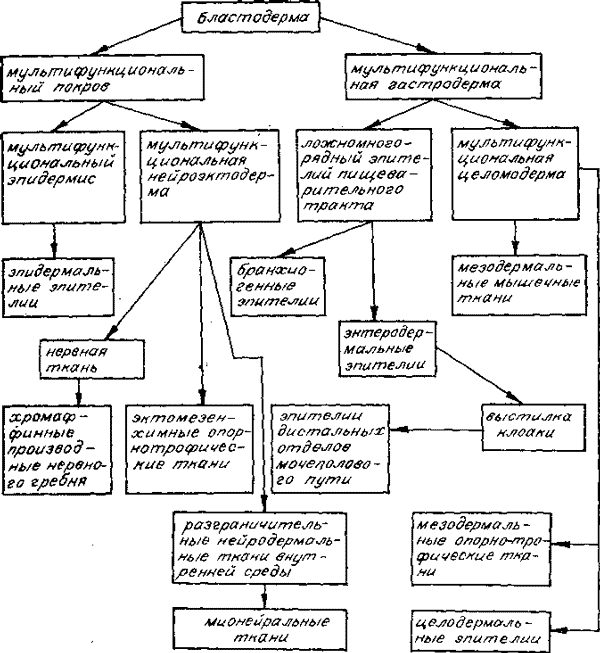
Филогенетическая преемственность тканей на пути эволюции к позвоночным.
Близость зачатков ларвальной мезодермы к нервной трубке рекапитулирует первично перибластопоральное положение протоцеломов. Напротив, изменения, происшедшие с бластопором: неполная рекапитуляция его смещения и совершенное выпадение в онтогенезе удвоения бластопора, сделали невозможным рекапитуляцию первичного способа образования хорды.
Рекапитуляционное значение последующих фаз онтогенеза позвоночных вызывает меньше разногласий и потому здесь не рассматривается.
Филогения тканей позвоночных показана на рисунке. Состав названных генетических групп тканей будет уточнен в следующем разделе.