Выстилка пищеварительной трубки ланцетника ложномногорядная, мерцательная
После стабильного переворота зона замыкания бластопора стала спинной, а с питанием возникли трудности, рот отделился от грунта (дорсальное положение приобрел и анус, что воспроизводится у личинок асцидий).
Далее возможна была смена объектов питания, переход от сбора ила к улавливанию планктона. Этот путь привел к асцидиям. Второй путь — изменение положения рта и сохранение питания илом. Этот путь вел к позвоночным. Есть, однако, указания, что рот позвоночных возник путем слияния двух передних жаберных щелей [Шмальгаузен, 1937]. В рамках рассмотренной модели это понятно. Смещение рта — процесс длительный, а питание требуется повседневно. Жаберные отверстия оказались ближе к грунту, чем рот, и это определило субституцию. Отсюда следует, что рту асцидий соответствует назогипофизарный канал круглоротых.
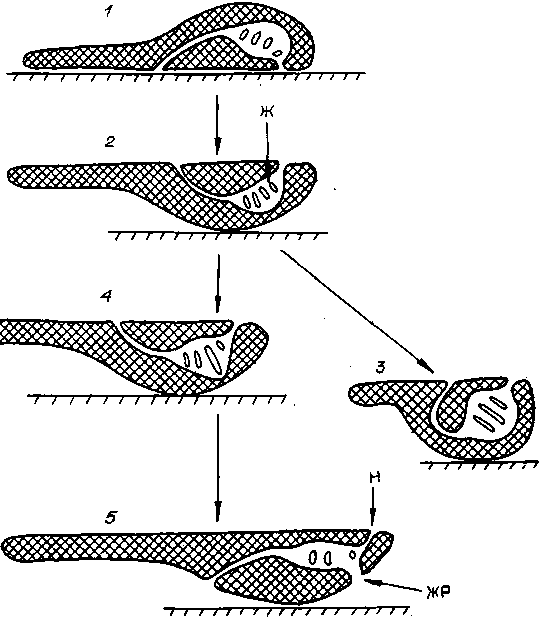
Схемы хордовых животных, вид сбоку.
1 — до превращения первично брюшной стороны в спинную, 2 — после этого, 3 — предок асцидий, 4, 5 — стадии превращения в цефалохлордовое животное. Ж — жаберная кишка, ЖР — рот, возникший слиянием двух жаберных щелей, Н — назогипофизарный канал.
Ввиду существенных гистологических различий полухордовых, оболочников, бесчерепных и позвоночных необходимо обсудить также возможные гистологические особенности предка позвоночных.
Выстилка пищеварительной трубки ланцетника ложномногорядная, мерцательная. По структуре пласта, наличию реснично-микровиллярных клеток, мукозных и белковых гландулоцитов она сходна с энтеродермой баланоглоссов. Эти черты поэтому можно считать древними. Однако нервных элементов в энтеродерме ланцетника нет, и в отличие от баланоглоссов эта выстилка— эпителий, а не существенно мультифункциональная ткань. Сходна с таковой у ланцетника энтодерма личинок миног. Поэтому можно считать, что такой эпителий был свойствен и провертебратам. Возник он из ткани типа энтодермы кишечнодышащих через исчезновение в ней нервных клеток.