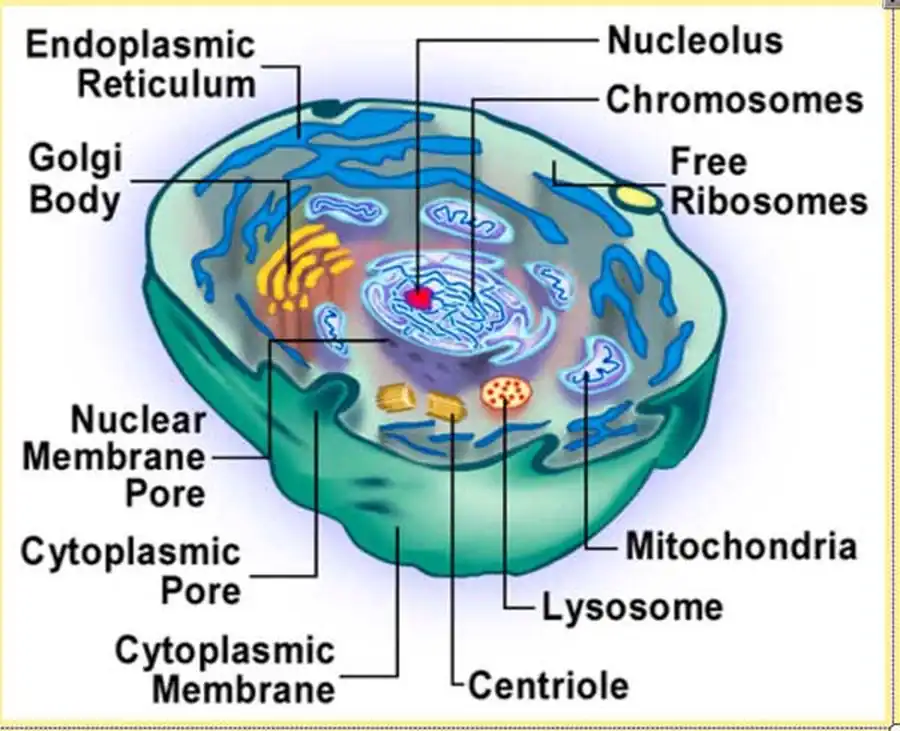
У ланцетника и позвоночных хорда состоит из совершенно разных тканей
К этому периоду надо отнести и возникновение хорды у общего предка полухордовых и хордовых.
У ланцетника и позвоночных хорда состоит из совершенно разных тканей [Welsch, Storch, 1973]. Это может означать, что при своем возникновении хорда состояла из очень примитивной мультифункциональной ткани, которая могла далее эволюционировать в разном направлении у различных вторичноротых.
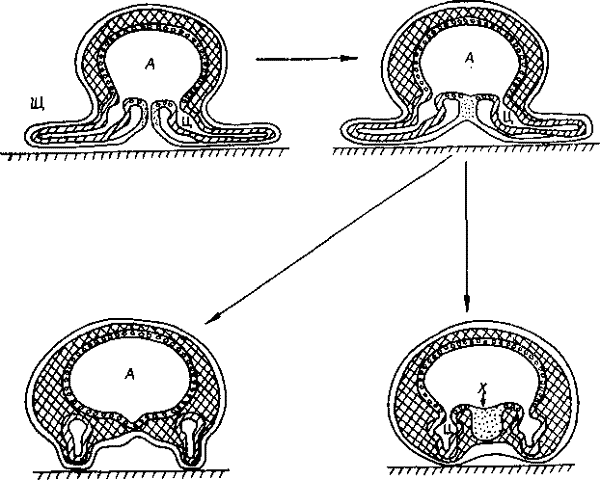
Поперечные срезы протоцеломического животного в период замыкания середины щелевидного бластопора.
А — архентерон, Д — протоцеломы, Щ — щупальцы, X — хорда.
Хорда лежит дорсально, но в ходе эволюции хордовые перевернулись (ниже), следовательно, первичное положение хорды — на стороне замыкания щелевидного первичного рта. И возникновение хорды могло быть связано с этим замыканием, которое в одном из вариантов в качестве побочного продукта дает хорду.
Если бы хорда возникала сразу для выполнения определенных функций, ее ткань у ланцетника и позвоночных обнаружила бы некоторое родство. Но у ланцетника она — своеобразная мышечная, а у позвоночных — тургорная. Это легко могло быть, если хорда возникла как побочный продукт замыкания бластопора, а функциональное значение (не совсем одинаковое у разных хордовых) она приобрела вторично.
Переворот хордовых так, что прежняя брюшная сторона стала у них в ходе эволюции спинной, предполагал еще Ж. Сент-Илер. Подробные и достаточные доказательства обобщены В. В. Малаховым [1977]. Отсюда, кстати, следует, что нервная трубка цефалохордовых развилась из циркумбластопорального сплетения, а ларвальные мезомеры закладываются в зоне вокруг первичного рта. Это подкрепляет предположение о том, что целомы хордовых — дериваты не гастроваскулярных каналов, а внутренней выстилки щупалец. Неясен, по мнению В. В. Малахова, лишь механизм переворачивания хордовых. Достаточно естественную модель, однако, можно предложить.