Первичные Metazoa представляли собой сплошную массу клеток
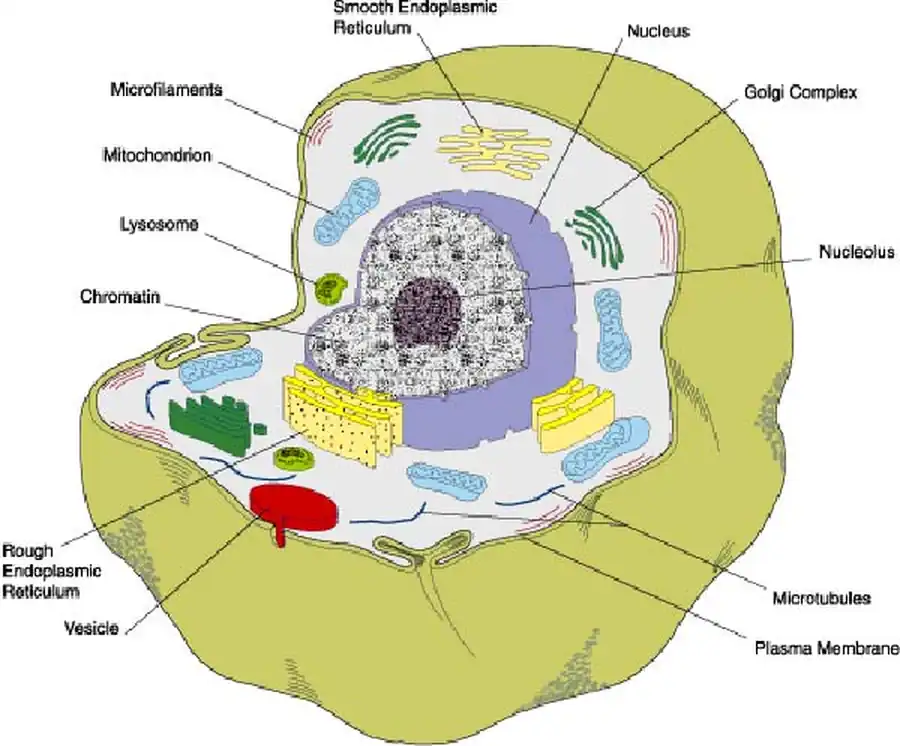
Предположение, что первичные Metazoa представляли собой сплошную массу клеток, высказывалось А. А. Заварзиным (1953) и В. Н. Беклемишевым [1964].
И хотя в экологическом плане эти модели не проработаны, гипотезу надо оценить. Вероятнейший предок таких животных — «радиальная» колония. При ее прогрессивной эволюции число клеток должно расти. Но при соединении клеток «конец в. конец» в крупной колонии, где клетки лежат тесно, при увеличении числа клеток неизбежна смена структуры колонии — или часть клеток потеряет контакт с поверхностью и возникнет однородная паренхимелла, либо все клетки сохранят контакт с поверхностью колонии, но перейдут на контакты «бок в бок» и возникнет полая сфера.
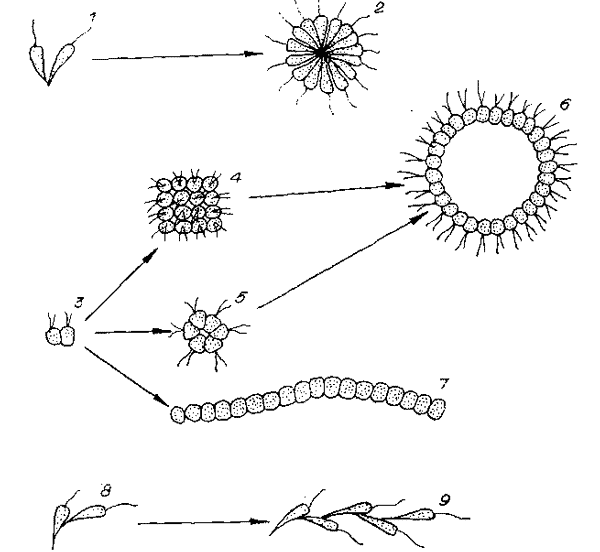
Зависимость морфологии и эволюции колоний от способа соединения клеток.
1 — соединение «конец в конец», 2 — схема «радиальной» колонии, 3 — соединение клеток «бок в бок», 4 — табличная колония, 5 —мелкая сферическая колония, 6 — бластееобразная колония, 7 — цепочка клеток «конец в бок», 8 — соединение «конец в бок», 9 — древовидная колония.
Для проверки этих возможностей производился искусственный отбор мутантов в культурах колониальной зоотрофной евглены Anthophysa vegetans [Борисов, 1978б]. В систематическом отношении евгленовые далеки от предполагаемых предков Metazoa (хоанофлагеллят). Однако изучавшиеся колонии очень сходны по организации с радиальными колониями хоанофлагеллят. Но если верно, что чисто морфологические особенности колонии существенны для ее эволюционных тенденций, то избранный объект вполне подходит. Его достоинство — быстрота размножения.
Эта евглена образует колонии, содержащие от нескольких штук до нескольких десятков клеток. Размножение бесполое путем деления колонии на равные или неравные части. При пересеве на свежую культуральную среду образуются очень мелкие колонии, которые быстро растут и выделяют слизистую нить. Нитью колония может прикрепляться к субстрату. При делении колонии нить разветвляется. С истощением среды колонии отделяются от нити и становятся бродяжками. При новом пассаже цикл повторяется.