У прокариот регуляторные участки ДНК невелики
Распространенные представления о том, что первоначально биосфера была представлена лишь прокариотами, а позже некоторые из них превратились в эукариот, возможно, и не верны [Reanney, 1974; Woese, Fox, 1977].
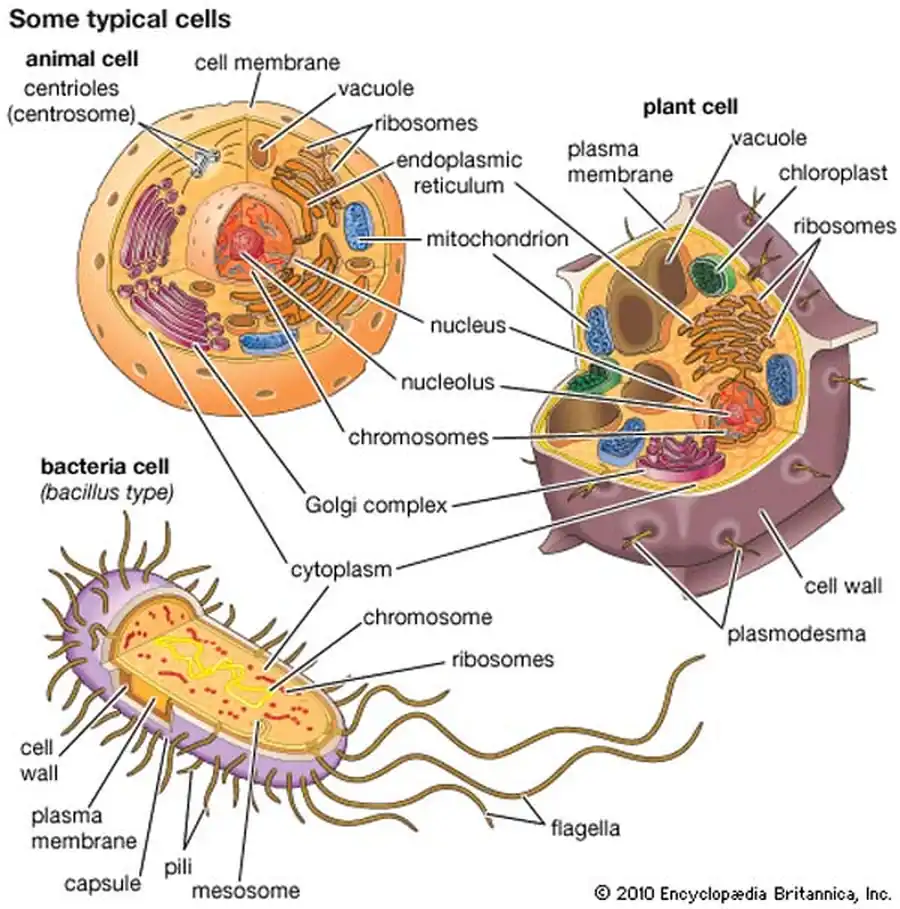
Принципиальные отличия генетического аппарата про- и эукариот [Дубинин, 1979] свидетельствуют о том, что это — расходящиеся ветви эволюции; и расхождение могло начаться прямо от протобионтов. У прокариот регуляторные участки ДНК невелики, преобладают структурные гены. Копии РНК, образующиеся на ДНК, прямо используются в синтезе белков.
У эукариот, напротив, велики отрезки ДНК, не кодирующие белков. Начало считыва ния ДНК-матриц, видимо, может происходить в разных точках (и, следовательно, разными способами, а не одним, как у прокариот). РНК-копии у эукариот подвергаются ферментативной обработке с «вырезыванием» части «текста» и сшивкой оставшихся участков. При этом только и образуется иРНК, готовая к трансляции. Вероятно, эта особенность и вызвала появление ядра, которое суть не столько хранилище генетической информации, сколько аппарат переработки первичных копий в иРНК.
Истоки различий про- и эукариот могут корениться в эволюции первичных полинуклеотидов. Возникли они абиогенно, и в них были неизбежны «бессмысленные» участки, не нужные для обеспечения жизнедеятельности протобионта. Один из путей преобразования таких молекул — делеция «бессмысленных» участков матриц. Такой путь выгоднее для мелких протобионтов, которые из-за малости объема не могут позволить себе никаких излишеств. Это — путь к прокариотам. Второй путь эволюции — сохранение «избыточности» в первичных матрицах и уничтожение излишних участков в копиях.
Трудно сказать, как могла возникнуть и совершенствоваться такая система, но она возникла. Впоследствии «избыточные» участки ДНК стали регуляторными. Второй путь эволюции труднее и длиннее. Но в итоге эукариоты получили преимущество — возможность по-разному считывать имеющуюся генетическую информацию (образно говоря, они могут из слова «гистология» составить слова «стол», «лог», а может быть и «гол»; прокариоты «читают» это слово только целиком). При этом равный отрезок ДНК в эукариотной клетке кодирует больше информации. А возможность считывания матриц разными способами предполагает наличие у всех эукариот фенотипического разнообразия.
«Многоликость» клетки заложена в организации ее генетического аппарата. Отбор фенокопий может закрепить несколько или один из возможных фенов как обязательный в жизненном цикле и «убрать» другие возможные фенотипы за счет перестройки системы генов-регуляторов. Но тенденция к разнообразию формы и обмена на основе того же генотипа останется как фундаментальное свойство эукариотной организации. Это — предпосылка к появлению сложных протозойных жизненных циклов со сменой форм клеток и одновременно «обещание» разнообразия клеток многоклеточных животных.